イメージギャラリー : フォトギャラリー
バイオミネラル
画像をクリックすると拡大して表示されます
1. 過飽和溶液より析出した炭酸カルシウム(CaCO3)の多形(SEM)

鉱物には、化学組成が同じであるがその原子配列(結晶構造)が異なるものがよく存在する。典型的な例は炭素のみでできたダイヤモンドと石墨(グラファイト)である。このようなものは同質多像あるいは多形と呼ばれる。 代表的なバイオミネラルである炭酸カルシウム(CaCO3)にもいくつかの多形が存在し、バイオミネラルとしては、カルサイト(方解石)、アラゴナイト(霰石)、それにファテライトと呼ばれる多形が見られる。写真は炭酸カルシウムの過飽和溶液から析出したこれらの多形で、(a)カルサイト、(b)アラゴナイト、(c)ファテライトである。 カルサイトの自形として、このように滑らかな結晶面を菱面体がよく見られる。またアラゴナイトは六角形の断面をもつ柱状あるいは針状の結晶が一般に発達する。ファテライトは板状や円盤状の結晶として見られることが多い。 バイオミネラルにおける炭酸カルシウムでは、このような自形が見られることもあるが、概して、これとはまったく異なる結晶の形となることが多い。
![]()
2. ザリガニの胃石を構成する非晶質炭酸カルシウム(SEM)

バイオミネラルを構成する炭酸カルシウムのほとんどは、カルサイトなどの結晶となっているが、ごくまれに結晶とならない(つまり規則的な原子配列をもたない)非晶質のものがあり、非晶質炭酸カルシウム(amorphous calcium carbonate: ACC)と呼ばれる。 アメリカザリガニの胃石(右の写真の白い玉のようなもの)は、その例のひとつである。この胃石はザリガニの脱皮前にカルシウムの貯蔵器官として形成されることが知られている。高分解能なSEM像(左)で見ると〜100 nm程度の顆粒状の物質が見られるが、これがACCで、それを結ぶ紐のようなものはキチン等の有機質であろう。 このような形状のACCは、適当な溶液の条件の変化によって容易に溶解し、脱皮後に形成される新しい外骨格(殻)に迅速にカルシウムを供給できると考えられる。
![]()
3. シロザケの星状石(SEM)

1.でバイオミネラルに見られる炭酸カルシウムの3つの多形を紹介したが、その中でもファテライトの出現頻度は他の2つの多形に比べ、かなり少ない。 魚類の耳石のうちで星状石と呼ばれているものは、ファテライトでできているバイオミネラルの数少ない例である。ただ我々が養殖ニジマスを調べたところ、耳石中の多形はそれほど厳密にコントロールされていないらしい。 写真はシロザケの星状石の低倍でのSEM像であるが、まさにミクロな枝サンゴのような特徴ある形状をしている。
![]()
4. 貝(軟体動物)の真珠層(SEM)

軟体動物の貝殻内側に形成される真珠層は、宝石の真珠や螺鈿の材料として見られるように美しい光沢を放ち、常に人々を魅了してきた。 その光沢は、左のように、光の波長と同等の厚さをもつアラゴナイトの板状結晶が、薄い有機膜を挟んできれいに積み重なっていることに起因する。その美しさ故か、バイオミネラルの中でこの真珠層は最も多くの研究がなされてきた試料のひとつである。しかし、未だにその形成機構にはわからないことも多い。 写真左は二枚貝のアコヤガイ(左上の写真でAの四角の部分)、右は巻貝のコシダカガンダラの真珠層(右上の写真で赤い四角で示した部分)のSEM写真であるが、二枚貝と巻貝では真珠層の形成プロセスが大きく異なることを示している。 巻貝に見られるこの構造は英語の論文では“Stack of coins”などと表現されているが、我々にはまさに“賽の河原”という表現がぴったりである。
![]()
5. エチレンジアミン四酢酸溶液でエッチングされたアコヤガイ稜柱層の表面(SEM)

アコヤガイの真珠層の外側には稜柱層と呼ばれる貝殻構造が形成されている。4.の真珠層がアラゴナイトで形成されているのに対して、この稜柱層は同じ炭酸カルシウムでもカルサイトでできている。 このSEM写真はアコヤガイの内側の、真珠層が形成された領域の外側に見られる稜柱層(4.の左上写真のBの四角部分)を、エチレンジアミン四酢酸(EDTA)の溶液でエッチングしたものである(右は左の写真を一部拡大)。 黒く見える枠組みは稜柱間壁と呼ばれる有機基質で、この中にカルサイトの結晶が形成される。興味深いことにEDTAでカルサイトをエッチングすると、非常に複雑な形状の“割れ目”が結晶の表面に現れてくる。解析の結果、この割れ目を境にしてカルサイトの結晶方位が多少異なっていることが明らかとなった。
![]()
6. コガモガイの交差板構造(SEM)

貝には真珠層、稜柱層以外にも興味深い微細構造がいろいろと見られる。 このSEM写真はカサガイの一種であるコガモガイの殻の内側を撮ったもので、この構造は“交差板構造”と呼ばれている。 この貝殻は5層の異なる構造が積層しており、その(外側から数えて)第3層と5層がこの構造を有する。薄い短冊状のアラゴナイト結晶が、斜めに折り重なっている様子は、ちょうど日本の古社寺の屋根に使われる「檜皮葺」によく似ている。さらに結晶の折り重なる方向が、この写真のように十数µmごとに反転している(“交差板”構造と呼ばれる所以)。 はたして生物はどのようにしてこのような複雑な構造をつくりあげているのか、説明に窮する。
![]()
7. ムラサキウニの棘(SEM)

ウニの殻や棘も炭酸カルシウムのカルサイトでできているが、貝殻などとは違ってかなりの量のマグネシウム(Mg)が結晶中のCaを置き換えている。 このSEM写真はムラサキウニの棘を次亜塩素酸(HClO)に浸漬して、有機物を完全に取り除いたもので、左が棘の断面の中心部分、右は柱(左右に伸びている)の表面を撮ったものである。 その断面は穴だらけで、また表面は複雑な形態をしているが、1本の棘はカルサイトの単結晶となっており、そのc軸が棘の方向と一致している。ただしこの結晶はかなりの量の有機成分を含み、このためその機械的性質が純粋なカルサイトと大きく異なっている。
![]()
8. 円石(ココリス)(SEM)

海洋プランクトンの一種である円石藻がその周囲に形成する円石(ココリス)も非常に特徴的な構造をもつバイオミネラルである。 この写真はPleurochrysis carteraeという種(全体の写真を左上に示す)のココリスであるが、そのリングはカルサイトでできた2種類のユニットで構成されている。 その2種類のユニットは、その形状と共に楕円面に対する結晶方位が大きく異なっており、どのようにしてそのようなコントロールができるのかは興味深い問題である。
![]()
9. アラゴナイト中の原子配列(TEM)

1939年に産声を上げた透過電子顕微鏡(TEM)の分解能は飛躍的に向上し、現在では日常的に結晶中の原子配列を直視することができる。 この写真は、TEMを用いてアコヤガイ真珠層のアラゴナイト結晶を高分解能で観察したものであり、そこに見える周期的なコントラストは、まさにアラゴナイトの原子配列に対応している。 同じ方向から見たアラゴナイトの結晶構造(左上)の中で、白い楕円で囲まれた原子配列のドメインが写真の黒い部分に対応する。
![]()
10. アコヤガイ幼生の初期貝殻(TEM)

貝は受精後、水中を遊泳しながら十数時間で胚の周囲に最初の貝殻(幼殻)を形成する。 >左下の写真は、受精後18時間のアコヤガイの幼生が初めて幼殻を形成したときのものであるが、この試料を固定し、その厚さ0.3µmの殻をTEMで観察したものが上の写真である。この結晶はアラゴナイトで、約60度に交差した直線的なコントラストがよく見える。 これをさらに拡大して見たものが、右下の写真である。これはアラゴナイト中によく見られる双晶という一種の欠陥構造であるが、このように高密度に双晶が導入された結晶は非常に珍しいものである。
![]()
11. アコヤガイ靱帯中の針状結晶(TEM)

二枚貝には、2つの殻の接合部に蝶番靱帯と呼ばれる組織が存在する。貝柱は貝を閉じる方向に働くが、この蝶番靱帯は逆に殻を開く方向に作用する。味噌汁の中のアサリが常に開いているのは、このためである。この蝶番靱帯には繊維状の結晶が含まれていることが知られている。 アコヤガイの靱帯(4.の左上の写真のCで示した部分)の断面をTEMで観察すると、まさに鉛筆を束ねて上から見たような構造となっている。六角形のひとつひとつがアラゴナイトの結晶であり、その間を有機基質が埋めている。各結晶でコントラストが違うのは、微妙な結晶方位の違いによるもので、本質的なことではない。 六角形の対角線のひとつに筋が見られるが、これは10.の写真にもあったアラゴナイトの双晶である(右下はそれを示す高分解能像)。 非常に揃ったサイズや、このように規則的に双晶が形成されるなど、バイオミネラルの不思議さを示す試料と言える。
![]()
12. 磁性細菌中のマグネトゾーム(TEM)
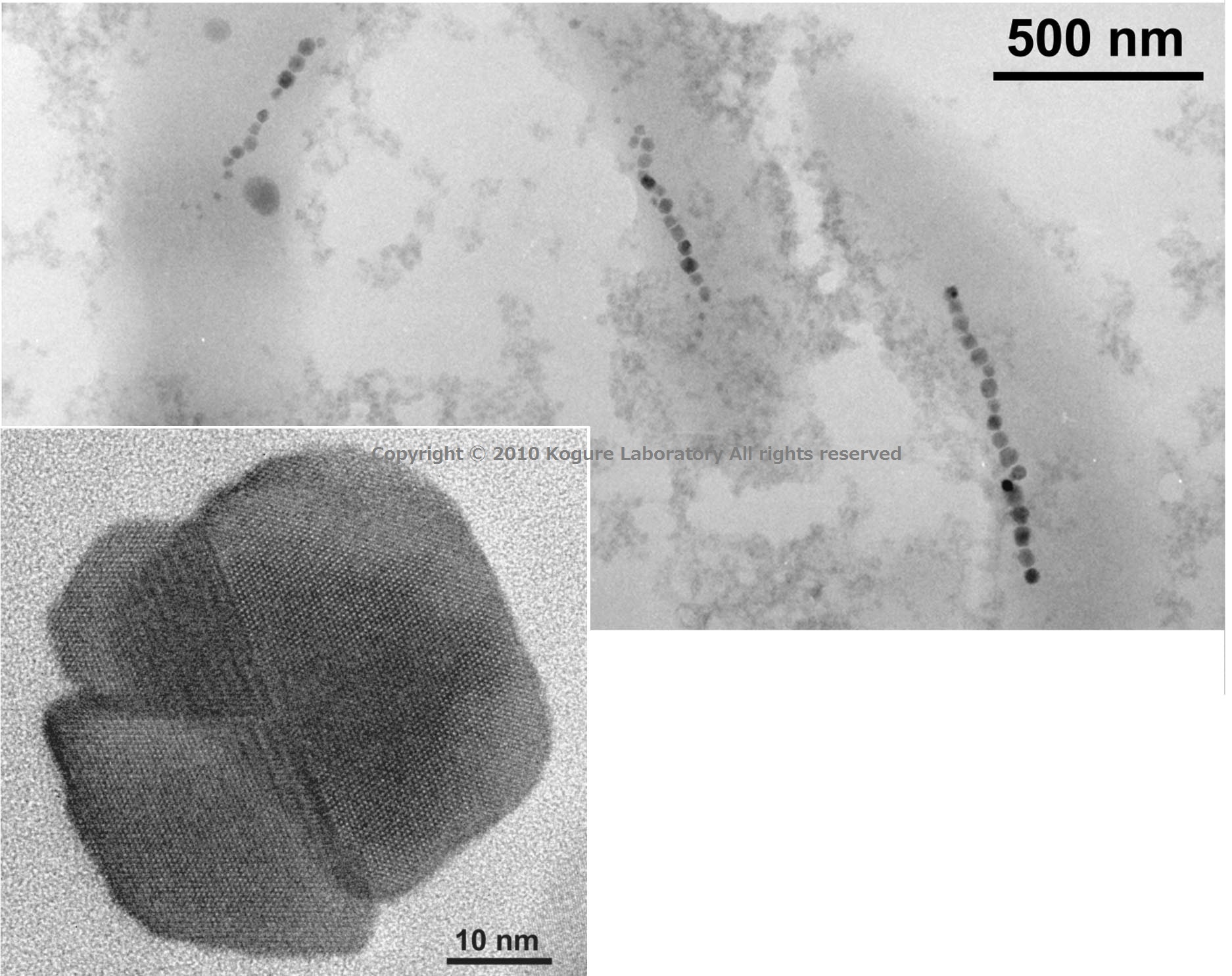
これまで紹介したバイオミネラルはすべて炭酸カルシウムであったが、この物質以外にも様々なバイオミネラルが報告されている。例えば骨や歯を形成するアパタイトや珪藻の殻をつくる非晶質シリカなども典型的なバイオミネラルである。 ここに示したのは、磁性細菌中のマグネトゾームと呼ばれる酸化鉄(Fe3O4)の微粒子である。その大きさは数十nmと小さいが、これはひとつの“磁石”となっていて、細菌にとって磁気センサーとして働く。 TEMで観察すると、アラゴナイトと同じく双晶がよく含まれていることがわかる。
![]()